Evolučná ontológia
Zdá sa, že vývoj v oblasti evolučných vied naznačuje na možnosť definovania evolučne orientovanej ontológie. V tomto cykle sú články, ktoré by mohli pripsieť k definovaniu evolučnej ontológie.
Podobnosť stavby mozgov vtákov a cicavcov kladie darwinizmu ťažké otázky.
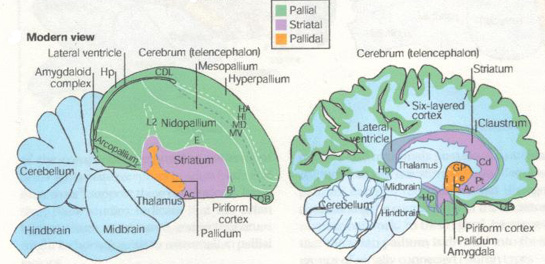 Hoci sa vývoj vtákov od plazov oddelil pred asi 150 mil. rokov a cicavcov pred asi 250 mil. rokov, mozgy vykazujú v nových štruktúrach (mimo tzv. plazí mozog) výraznú architektonickú podobnosť, aspoň podľa [1].
Hoci sa vývoj vtákov od plazov oddelil pred asi 150 mil. rokov a cicavcov pred asi 250 mil. rokov, mozgy vykazujú v nových štruktúrach (mimo tzv. plazí mozog) výraznú architektonickú podobnosť, aspoň podľa [1].
Na prvý pohľad by sa mohlo zdať, že zrejme museli riešiť rovnaké problémy, a to vyvolávalo selekčný tlak na vývoj rovnakých štruktúr. Veď aj inde vidíme podobné príklady, kto chce lietať musí mať krídla, kto chce plávať musí mať plutvy. Napokon autori predloženej štúdie, vychádzajúc z rovnakej myšlienky si robia nádeje na nájdenie akýchsi obecných princípov myslenia, a tie by radi uplatnili v počítačoch.
Pravdou však je, že napríklad plutvy kytovcov vznikli zrastením zadných končatín, kdežto plutvy rýb sformovaním zadnej chrbtovej časti tela. A konštrukcia krídla vtáka a hmyzu je niečo architektonicky zásadne iné. Prečo teda na riešenie rovnakých úloh mozgu treba rovnakú architektúru?
Druhá možnosť je, že zákonitosti neskoršej stavby mozgu sú už v značnej miere zabudované do zákonitostí predchádzajúcej stavby. Teda z toho, akú architektúru a funkcionalitu má plazí mozog, vyplýva jediná možná architektúra neskorších mozgov. Pravda za darwinisticky predpokladaného rovnakého selekčného tlaku prostredia, teda, že funkcie mozgu sú dané potrebami z prostredia. Zvláštne je aj to, že mozog článkonožcov tiež vykazuje určité zhody s mozgami stavovcov, aj keď sa vyvíjali odlišne, viď [2], samozrejme na oveľa základnejšej úrovni.
Ak by tomu tak bolo, treba zrejme prinajmenšom obmedziť predstavu náhodných zmien, ktoré by mali byť aj podľa dnešného syntetického darwinizmu zdrojom evolučných noviniek. Vývoj by potom vyzeral skutočne teleologicky, teda akoby bol účelovo ťahaný určitým smerom. Ovšem potom sa nedá ubrániť otázke zámeru.
Náhodnosť zmien, ktorú predpokladá napríklad génocentrický model evolúcie je vo svojej podstate tautológia. Ak predpokladáme že gény sú agenti, ktorí sa neustále snažia zachovať svoju štruktúru presným replikovaním, tak podstata diania je v replikácii. Z toho samozrejme nemôžu vznikať nijaké evolučné novinky a diverzita, na ktorej by sa realizoval prírodný výber. Zdroj zmien potom musí vychádzať z javov, ktoré nie sú javovou podstatou replikácie, a kedže nie sú ani javovou podstatou prírodného výberu, tak sa zvalia na nepochopiteľnú a neuchopiteľnú náhodu. Zmena sa stane tak, že sa niečo zmení a hotovo. Náhoda tu robí zástupcu Božieho zázraku, pretože zázrak je zmena, ktorá nepochádza zvnútra sveta, ale zvonka, od Boha.
Lenže ako spadá do toho to, že mozgy (a nielen mozgy) sa u rozličných druhov od seba evolučne vzdialených vyvíjajú rovnakým smerom, hoci náhodná zmena by ich mala posúvať k rôznym riešeniam tých istých problémov? Žeby sme miesto Božieho zázraku pre zmenu mali neustálu prítomnosť Božieho prstu?
Podľa mojej mienky sa dá neustálej Božej prítomnosti (samozrejme nie stvoreniu Sveta Stvoriteľom) vyhnúť tak, že vylúčime náhodu úplne. Predpokladáme potom svet prísne deterministický od počiatku doteraz. V takom svete je na počiatku dané určité usporiadanie (kto chce nech si verí, že je to Slovo), kde každý objekt sveta vytvára súčasne seba a súčasne aj svet, teda aj zákonitosti, ktoré v takom svete platia. V takomto svete je stav súčasných usporiadaní objektov určujúci pre každý následný krok evolúcie, pretože jednak vytvára štruktúry toho, na čo sa uplatnia budúce zákonitosti sveta, teda samotné objekty, ale aj štruktúru tých budúcich zákonitostí, ktorými sa budú musieť objekty riadiť. Nebolo by teda náhodné, ako čo bude vyzerať o milión rokov, alebo miliardu.
Myslím, že týmto by sme mohli vyvarovať aj mystiky účelnosti teleologickej alebo teleonomickej. Účel totiž predpokladá cieľ, či už tak alebo onak. Nepomôže problém ofintiť tak, že účel síce nie je v nejakom cieli, ale iba v smere vývoja. V oboch prípadoch sa neobídeme bez zámeru, a ten by sme museli predpokladať u Boha. Ak ho nechceme, neostáva iné ako vyššie popísaná predstava súčasnej evolúcie objektov sveta a zákonitostí sveta, pričom nič nestojí nad ničím a nič nie je svetu dané zvonka.
- Shanahan, M., Bingman, V. P., Shimizu, T., Wild, M., & Güntürkün, O. (2013). Large-scale network organization in the avian forebrain: a connectivity matrix and theoretical analysis. Frontiers in Computational Neuroscience, 7(July), 1–17. doi:10.3389/fncom.2013.00089. http://www.frontiersin.org/Computational_Neuroscience/10.3389/fncom.2013.00089/full
Neustála premena tvaru
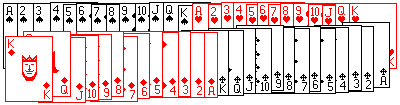
|
| Obr. 1. Usporiadaný balíček kariet A |
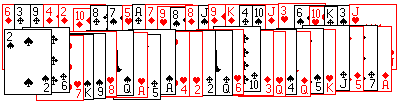
|
| Obr. 2. Neusporiadaný balíček kariet B |
Predstavme si dva balíčky kariet. V balíčku A (obr 1) sú karty usporiadané takto: najprv ♠ vzostupne od esa po kráľa, pod tým rovnako usporiadané ♣ , ♥ a napokon ♦ . V balíčku B (obr. 2) sú karty úplne náhodne premiešané. Položíme si nasledovné otázky:
- Ktoré z usporiadaní (A alebo B)
je pravdepodobnejšie?
- Ktoré z usporiadaní (A alebo B) obsahuje viac informácie?
Nemyslime teraz na to, že z pohľadu fyziky ide v podstate o tú istú otázku, len inak položenú. Povedzme, že karty budeme usporiadavať tak, že ich nasypeme na stôl, a potom zhrnieme do balíčka. Aká je pravdepodobnosť, že pri jednom pokuse vznikne balíček A a aká, že vznikne balíček B? Že nastane akékoľvek náhodné usporiadanie (prípad B) je prakticky rovná jednej, teda P(B) = 1. Aby nastalo konkrétne usporiadanie A treba neskutočný počet pokusov (ak som správne počítal tak je to 67-miestne číslo 8.066x1067), teda pravdepodobnosť pri jedinom pokuse je skoro nulová,
P(A) = 1.24x10-68
.
Chce sa tiež povedať, že balíček A obsahuje viac informácie ako balíček B, pretože okrem toho, že obsahuje skupinu symbolov hodnôt (informácie o hodnote), obsahuje aj informáciu o ich následnosti.
Za takýmto riešením je však mlčky predpokladané, že balíček kariet nie je sám o sebe, že nie je jedno, ktoré usporiadanie symbolov je správnejšie, ako iné. Ak by sme predložili takúto úlohu niekomu, kto nikdy žiadne karty nevidel, nepozná čísla ani symboly, asi by ťažšie rozoznal, v ktorom balíčku je aké usporiadanie a jeho odpoveď by bola, že obe riešenia sú rovnocenné, teda A=B. Podobne, ako nám môže pripadať rovnocenné akékoľvek náhodné zamiešanie kariet.
Balíček kariet sám o sebe je nesporne nejako usporiadaný, obsahuje informáciu sám o sebe a obsahuje jej rovnaké množstvo, nech je usporiadaný akokoľvek. Taktiež je pravdepodobnosť každého z týchto usporiadaní rovnaká.
Možno si čitateľ všimol, že aj na tomto jednoduchom príklade som v zastúpení používal slová usporiadanie a informácia. Naozaj, som presvedčený, že sú to dve slová, ktorá označujú skutočnosť, že prvky sú nejako voči sebe uložené, či už v priestore, čase, alebo prípadne inak. Ak po srdcovom ese nasleduje kárová osmička ide o inú informáciu, ako keby tomu bolo naopak, ide o iné usporiadanie.
Kým sa nepoužívalo slovo informácia, myslitelia používali vzťah formy a obsahu. Chápali ich nedeliteľnosť. Ak vezmem hlinu a vymodelujem z nej misku, je tu hlina a z nej forma — miska. Ak z toho istého množstva hliny vymodelujem džbán, je tu tá istá hlina, ale forma — džbán. Ten istý obsah sa objavuje v dvoch usporiadaniach, v dvoch formách, alebo iným slovom: obsahuje inú informáciu. Platón z toho dokonca vyvodil svoj systém ideí, kde chápal obsah ako nemenný, menila sa iba jeho forma, ktorú chápal ako myšlienku (eidos).
Slovo informácia vzniklo z tohoto konceptu niekedy v dvanástom storočí (aspoň v anglickom prostredí), citát z roku 1387: Päť kníh prišlo z nebies, aby in-formovalo (utváralo) ľudstvo. Božská idea usporiada do tvaru veľkého plánu ľudstvo. Nie je podstatný človek, nie je podstatné telo, to je iba obsah, ktorý Božia ruka formuje do správneho usporiadania. Správnosť toho je v Bohu, nie v ľudstve, až Božím aktom formovania nastane nová in-formácia.
Nie je účelom tohoto článku opisovať vývoj chápania slova
informácia. Dnes sa chápe rôzne. Vo fyzike ako opak neusporiadanosti, v
informatike ako správa, niekde ako akt premeny usporiadania... Za
všetkými definíciami však vždy je niečo, čo súvisí s usporiadaním. S
formou. Myslím, že nespravím nijakú chybu, ak budem v tomto texte, ale
aj v iných projektu Anima Et Machina, chápať informáciu ako
usporiadanie.
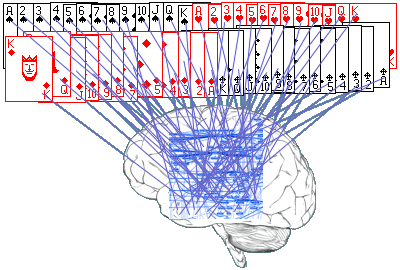
|
| Obr. 3. Kontext usporiadania kariet |
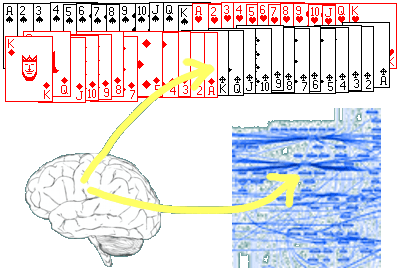
Vrátim sa k môjmu kartovému príkladu. Nech karty usporiadam akokoľvek, vždy obsahujú nejaké usporiadanie. Prečo práve je usporiadanie A výnimočné? Pretože je dané do vzťahu, kontextu, s kultúrou spoločnosti, ktorá karty používa. To, či je usporiadanie také, alebo onaké sa odohráva nie v tom balíčku kariet, ale v niečej hlave (obr. 3). Je to v konečnom dôsledku nejaké usporiadanie neurónov v mozgu (znázornené ako modrá štruktúra), ktoré sa vzťahuje (vytvára kontext) k jenotlivým symbolom na kartách.
Obrázok sa pokúša znázorniť nasledovnú predstavu. Balíček kariet je jedna informácia, štruktúra neurónov v mozgu je iná informácia. Pripomínam, že pod informáciou chápem usporiadanie. Kontext vzniká spojením dvoch usporiadaní v jedno. Karty spolu s neuronálnou štruktúrou v mozgu tvoria jedno usporiadanie, jednu informáciu.
Táto, na prvý pohľad samozrejmá konštrukcia, má závažný dôsledok. To, či nejaké usporiadanie je v kontexte s usporiadaním iným je čisto subjektívna konštrukcia. Máme sklon chápať usporiadanie kariet a usporiadanie v mozgu ako od seba nezávislé entity, ktoré vstupujú do kontextu. Ak by to tak bolo, zostáva zodpovedať otázku, ako to vôbec môžu spraviť? Veď karty a mozog, ak sú úplne nezávislé, potrebujú nejaký vzťah, ktorý ich spojenie vytvorí. Čo to je a odkiaľ sa to vezme? Existuje niečo mimo akúkoľvek informáciu, nejaké pole usporiadaní, ktoré vytvára kontexty? Niečo, ako kontext kontextov? Existuje nejaká superinformácia, Boh, niečo, čo tvorí kontext všetkého? Alebo slávne Sheldrakove morfogenetické polia?
|
| Obr. 4. Vznik kontextu |
Ak aj vychádzame z podobných predstáv, problémom zostáva vyčlenenosť, externalita takýchto polí kontextov voči informáciám, ktoré vzájomne dávajú do kontextu. Nemáme nijaký spôsob, ktorým by sme vedeli povedať, ako to ten Boh robí, že nás formuje, ako morfogenetické polia vlastne formujú orgsnizmy, pretože je to mimo nás, ako objekty toho procesu.
Napokon - veď nejde o nič iné ako usporiadanie usporiadaní do nového usporiadania. Prečo teda hľadať niečo iné, ako to, čo už je? Hľadajme opäť iba spôsob, ako sa usporiadania usporiadajú. Ani karty, ani predstava poradia sa predsa neobjavili z ničoho nič, akoby zázrakom. Majú svoju minulosť. Voľakedy niekto vymyslel kartové symboly, niekto vymyslel karty. A rovnako niekto (v tomto prípade ja) vymyslel, že ich usporiadanie v prípade A je význačnejšie, ako iné usporiadania (obr. 4). A ani ja som sa tu neobjavil zázrakom, rovnako som výsledkom súčasných a predchádzajúcich usporiadaní.
Ak sa pozrieme na vec z takého uhlu pohľadu, tak vlastne niet kontextu. Je iba jedno jediné usporiadanie, ktoré sa nestále v čase a priestore mení. Vytváranie kontextu je iba pomôcka pre pochopenie nedeliteľnej jednoty bytia. Vytváranie kontextu je nástroj, ktorým mozog modeluje prostredie, ktorého je súčasťou, ktorým vytvára obraz toho prostredia. To môže robiť iba vyčleňovaním z celku.
 |
| Obr. 5. Na počiatku bolo slovo |
Ak pochopíme usporiadanie, ako v čase sa meniacu, seba formujúcu štruktúru pochopíme, že evolúcia je iba iné slovo pre to isté. Evolúciu môžeme chápať ako zmenu usporiadania Vesmíru v čase. Dnešná predstava kozmológov o Vesmíre je taká, že vznikol z bodu s nesmiernou hustotou a teplotou procesom, ktorý nazývajú Big Bang. Z hľadiska našej témy je zrejmá predstava, že usporiadanie muselo byť už od počiatku. A skutočne, od samotného vzniku Vesmíru tu je. Kozmológovia nijako nevedeli zdôvodniť, ako mohli vzniknúť hviezdy, galaxie, ako vôbec vznikli akékoľvek štruktúry. A podľa mňa to ani nebudú vedieť vysvetliť. Veď si pokojne môžeme predstaviť Vesmír, ktorý vznikne podľa teórie Big Bangu, rozpína sa, chladne v každom svojom bode rovnomerne, hustota energie rovnomerne klesá. Prečo by vôbec malo v nejakom mieste vzniknúť akékoľvek zhustenie, alebo iná nerovnomernosť, akékoľvek usporiadanie? Vesmír by bol bez štruktúr. Ale nie je, inak by som to tu nepísal. Nerovnomernosti museli byť od saméhom počiatku, od momentu vzniku, museli byť súčasťou toho momentu, alebo mu predchádzať. A skutočne. pomocou mapovania tzv. reliktného žiarenia (obr. 5), teda zbytku z tej obrovskej počiatočnej energie Big Bangu, sa ukázalo, že od samého počiatku je Vesmír usporiadaný do štruktúr, od samého počiatku Vesmír obsahuje informáciu, teda že - na počiatku bolo slovo...
Niet iného kontextu, ako je celý Vesmír. Niet vyčlenenosti a nových entít, všetko je neustála premena tvaru, pôvodného usporiadania, ktoré si vytvára neustále podmienky pre nové preusporiadanie. V tom je podstata determinizmu bytia, osudovosti každého vyčleneného kontextu usporiadania.
Ako som už uviedol, zdá sa mi, že pojmy usporiadanie a informácie sa často používajú ako synonymá. Z axiómy jednoty a nedeliteľnosti bytia vyplýva, že každé vyčlenenie do pojmu, každý pojem, je iná časť bytia, ako tá časť, ktorú by pojem mal reprezentovať. Využijem preto príležitosť a skúsim pomocou zámenných pojmov popisovať iné aspekty toho istého. Je namieste zadefinovať si niektoré pojmy.
Replikátor pána Dawkinsa
Rozprava o evolúcii je vždy v princípe rozprava o opakovaní toho, čo interpretujeme pojmom informácia. Či už ide o organizmy, myšlienky, konformácie íľových sedimentov, skrátka akékoľvek usporiadanie, ktoré identifikujeme v rovnakosti formy, usporiadenie informácie v rovnakej konkrétnej in-formácii.
Vysvetlenie rovnakosti, ktorá núti zaraďovať organizmy do jednotlivých druhov a napokon vysvetlenie faktu, že jednotlivé druhy vo svojich znakoch na seba naväzujú spôsobili a dodnes spôsobujú jednu z najväčších intelektuálnych výziev pre ľudstvo. Možno sa dnes zdá, že darwinizmus (hlavne v jeho dnešnej post-neo-darwinistickej podobe) tieto otázky dokáže zodpovedať, ale všetko nasvedčuje tomu, že opak je pravdou.
Iste, dnešný model evolúcie, ktorý zredukoval pôvod rovnakosti foriem organizmov na rovnakosť v DNA, akoby dával víťazoslávnu odpoveď na základné otázky nášho bytia: kto sme a odkiaľ pochádzame. Faktom však je, že všetko vyzerá pekne pokiaľ sa nesiahne na základné predpoklady modelu.
Dnes prevládajúca evolučne-biologická paradigma (tiež sa zvykne označovať slovom génostredná ) je postavená na myšlienke, že jednotkou prírodného výberu nie je druh, ani jednotlivý organizmus, ako si predstavoval Darwin, ale gén, teda každý jednotlivý znak organizmu. Znakom je napríklad farba kvetu, alebo dĺžka chodidla, či schopnosť sa učiť. Navyše post-neo-darwinistická predstava lokalizovala znak (gén) do molekuly DNA ako jej identifikovateľnú časť, ako sériu písmen, ktoré sa od potomka, k potomkovi neustále opakujú a zabezpečujú vytvorenie rovnakého znaku pre organizmus. Prírodný výber sa potom realizuje primárne na jednotlivých variantách častí DNA molekuly. Ak variant génu podporuje prežitie organizmu ako celku, tak sa opakuje v ďalších a ďalších potomkoch. Ak naopak znižuje schopnosť prežitia, tak je postupne prírodným výberom z populácie vyradený. Organizmus samotný je potom iba nástrojom na prežitie týchto génov, tzv. vehiklom (vehicle) , ako to postuloval Richard Dawkins v knihe Sebecký gén .
Žiaľ, táto jednoduchá a príťažlivá predstava evolúcie živých systémov pravdpodobne neobstojí. Dnes je už zrejmé, že prípad, keď jeden gén v DNA kóduje jeden znak organizmu je skôr výnimka ako pravidlo. Navyše do procesu vytvárania znakov organizmu nevstupujú iba samotné prepisy génov, ale do samotného prepisu vstupuje rad iných faktorov v organizme, ktoré samé o sebe môžu byť predmetom prírodného výberu (viď napríklad A.Markoš Povstávání živého tvaru ). Dnes je skoro isté, že predstava Crichtonovho Jurského parku , kde vložíme do vajíčka vtáka DNA dinosaura a vznikne vyhynutý jašter je neuskutočniteľná. Ako hovorí Markoš: „je možné, že týmto postupom vznikne nepodarený vták a nie dinosaurus.“
Projekt Anima Et Machina však nie je primárne o biológii a ja sa necítim byť povolaný k tomu, aby som posudzoval jednotlivé biologické teórie. Koho zaujíma súčasný stav v tejto oblasti, môže si prečítať analytický a do podstaty veci idúci pohľad filozofa a bunečného biológa Antona Markoša v jeho knihe Tajemství hladiny . Mňa zaujíma iba jediný aspekt problematiky a tým je otázka podstaty evolúcie v biologickej oblasti, a to tým viac, že jeden z autorov súčasného génostredného pohľadu na evolúciu preniesol túto podstatu aj do oblasti ľudskej kultúry a dal tak základ myšlienkovému smeru zvanému memetika , teda oblasti, o ktorej môj projekt je.
V darwinistickej koncepcii evolúcie organizmy sú rovnaké v rámci jedného druhu s určitou mierou rovnakosti, a teda aj s určitou mierou odliškosti. Odlišnosť je podmienkou fungovania princípu prírodného výberu. Ak sú jedinci druhu málinko rozdielni, a neustále sa meniaca situácia v prostredí kde žijú vytvára pre určitú odlišnosť výhodu a pre inú odlišnosť nevýhodu, tak je vytvorený základný predpoklad pre selekciu, výber jedincov s výhodnými vlastnosťami pre ďalšie generácie. Postupné hromadenie malých zmien je pôdou pre vznik nových druhov a pre vznik neobyčajnej bohatosti foriem života na Zemi. Potiaľ veľmi zjednodušená predstava základných princípov darwinizmu.
Kľúčom k vysvetleniu akejkoľvek darwinistej predstavy teda je otázka: čo je podstatou a obsahom odlišnosti organizmov, teda na čom sa realizuje prírodný výber. No a tu vidíme v histórii darwinizmu zreteľný pohyb od druhu, cez jedinca druhu až po gén v molekule DNA. Richard Dawkins, o ktorom v tomto texte bude ešte veľa počuť, koncepciu evolúcie založenej na selekcii na úrovni génu v DNA obšírne obhájil a spopularizoval v knihe Sebecký gén . Ako evolučne zmýšľajúci vedec, ktorý musel v sebe prejsť cestu od prírodného výberu na úrovni druhu, na úrovni jedinca až po prírodný výber na úrovni génu v DNA, nevyhnutne narazil na otázku: odkiaľ sa zobrala DNA a čo jej podstatou.
Vzhľadom na to, že DNA je jedna obrovská molekula, tak nebolo ďaleko od myšlienky, že podstatou DNA nie je ani tak to, že je fyzikálne zložená z malých molekúl, ale to, ako je zložená, teda že nie je postatná fyzikálna vlastnosť molekuly, ale jej usporiadanie — informácia v nej uložená. Prírodný výber sa teda neuplatňuje na „hmotu“ DNA, ale na gény, chápané ako určité informačné jednotky. Jednotlivý gén je poradím znakov, písmen textu, podľa ktorého sa tvoria biologické štruktúry. Ani samotné štruktúry nie sú vlastne „živé“, „živá“ je informácia obsiahnutá v DNA. Ak sa v géne uloženom v DNA zmení jedno písmeno, zmení sa podľa tohoto génu produkovaná biologická štruktúra, teda konkrétna proteínová molekula.
Hoci prírodný výber pôsobí až na celý organizmus, teda produkt všetkých génov v ňom obsiahnutých, varianty génov v potomstvách zabezpečujú variabilitu organizmov, a teda pole pre prírodný výber. V konečnom dôsledku sa úspešná informácia v DNA presadí alebo nepresadí prostredníctvom celého organizmu.
Ak je teda podstatou jednotlivá informácia, tak nie je ani tak podstatný jej nosič, ale jej podstatné a určujúce vlastnosti: replikovateľnosť (opakovateľnosť) a variabilita . Replikovateľnosť zabezpečuje stabilitu informácie a viariabilita jej evolúciu. Na základe týchto predpokladaných vlastností molekulárneho génu vcelku logicky dospel Dawkins k predstave replikátora , teda nejakej entity, schopnej udržať informáciu tým spôsobom, že sa neustále rozmnožuje v ďalších a ďalšich generáciách, pričom pri rozmnožení nastávajú chyby, ktoré sú zdrojom variability a tým aj evolúcie. Aby zdôraznil „čistú“ nehmotnosť replikátora, v záverečných kapitolách knihy sa pokúsil nájsť iný typ replikátora. Našiel ho v ľudskej kultúre, kde jednotlivé myšlienky, alebo koncepty, návody, či iné artefakty sa replikujú z jednej hlavy do druhej, teda sú to informačné jednotky, vykazujúce obe vlastnosti: replikovateľnosť a variabilitu. Takéto jednotky nazval mimem skrátene mém , aby zdôraznil ich podobu s génami.
Dawkins takto priviedol na svet pojem replikátora. Z
epistemologického hľadiska by zavedenie tak podstatného pojmu vyžadovalo
jeho dôkladný rozbor a tiež dôkladné zdôvodnenie. Obávam sa, že toto
Dawkins neurobil, čo sa pokúsim v ďalšom texte doložiť. Pojem
replikátora obstal na intuitivnej úrovni, s čím sa Dawkins a mnoho
biológov a memetikov očividne uspokojilo a domnievam sa, že hlavne pre
memetiku, ako rodiacu sa vedu o ľudskej kultúre, to spôsobilo celý rad
koncepčných nedorozumení.
Tak ako — aktívne, či pasívne?
R.Dawkins pochopiteľne vie, že DNA je príliš zložitá molekula, aby vznikla nejakou náhodou. Preto predpokladá, že je produktom dlhého vývinu nejakej jednoduchšej molekuly, ktorej pravdepodobnosť vzniku je oveľa vyššia.
Predstava môže byť napríklad takáto ( Rozprávka o vzniku molekulárneho replikátora) :
Pred štyrmi miliardami rokov na Zemi vládla hustá atmosféra, plná uhľovodíkov. Existoval praoceán, a mohutná tektonická a sopečná činnosť. Tak oceán, ako aj atmosféra boli prehustené energiou. Neustále vznikali a zanikali chemické zlúčeniny s rôznou dobou rozpadu. Udržanie zložitejších molekúl v prostredí energeticky silných nárazov bol prakticky vylúčený. Napriek tomu neustále práve v týchto zrážkach vznikali a okamžite v nasledujúcej zrážke zanikali molekuly väčšie, ako sú základné zlúčeniny. Ďalej citujme autora:
Akonáhle takáto molekula vznikla, dôležité bolo, aby sa informácia, ktorá tvorí štruktúru replikátora preniesla do vytvorenia pokiaľ možno najväčšieho počtu potomkov skôr, ako sa materská molekula rozpadne. Z hľadiska stability informácie preto prestal byť dôležitý nosič informácie, ale rýchlosť a presnosť jej replikácie do ďalších a ďalších nosičov.
Kopírovanie ale občas nebolo presné a novo vzniknutí potomkovania nemuseli byť identickí. V tom momente začal fungovať princíp prírodného výberu. Ten potomok, ktorý sa dokázal rozmnožovať rýchlejšie ako iní jeho súrodenci, postupne v prapolievke na ich úkor prevládol, pretože vyčerpal zdroje (voľné chemické látky). Ten potomok, ktorý vydržal dlhšie vlastný rozpad, tak isto mohol mať viac potomkov. Úspešné zmeny sa hromadili, neúspešné vyhynuli. A tak sa mohol začať proces, ktorý postupne viedol k DNA a trvá v biologickej evolúcii dodnes.
Potiaľ rozprávka. Je natoľko príťažlivá, že je pomerne ťažké proti nej niečo namietať. Pravdou však je, že ju možno sotva dokázať, pretože stopy takéhoto procesu už dávno boli v evolučnom procese nespočetnekrát recyklované. Príťažlivosť pre evolučnú teóriu je najmä v tom, že pomerne logicky vidí DNA ako výsledok, nie začiatok evolúcie a DNA je molekula, teda musí byť dôsledkom molekulárneho vývoja.
Ako si Dawkins vlastne predstavuje fungovanie samotného replikátora?
V prvom rade sa mi vidí pozoruhodné, ak autor najprv uvedie príklad „ako to je mysticky“, teda toho Ezechiela, a hneď za tým čáry-máry a máme tu schopnosť sebareplikácie , ako dôsledok fyzikálnych zákonov. Nie som síce znalec fyziky, ale trúfam si povedať, že o sebareplikácii v nej nie je ani slova. Skrátka R. Dawkins čosi implicitne predpokladá a to „čosi“ je veľmi podstatnou vlastnosťou fyzikálnych objektov, čo je zároveň v istom zmysle nefyzikálna vlastnosť, pretože niet pre ňu fyzikálnych dôvodov.
Ako v citáte prvom, aj v tomto vidíme replikátor ako molekulu, ktorá sama tvorí svoje kópie. To značí, že je nadaná istou intencionalitou, schopnosťou aktívneho konania. Má informácie o prostredí a sama aktívne vyhľadá to, čo pre jej replikáciu potrebné je a vylúči to, čo pre jej replikáciu potrebné nie je. Vie sa rozhodovať . Napokon túto vlastnosť aj sám autor explicitne predpokladá:
Máme tu myšlienkovú konštrukciu: hypotetické vlastnosti niektorých replikátorov majú vplyv na pravdepodobnosť svojej vlastnej replikácie (moc nad vlastným osudom), čo je dôsledok sebareplikácie. Nuž aspoň v nejakej rudimentárnej podobe to pripomína draka Ouroborosa, ktorý požiera sám seba: proces replikácie prebieha tak, že replikátor má také pozoruhodé vlastnosti (schopnosť sebareplikácie, moc nad vlastným osudom), ktoré mu umožňujú proces replikácie. Obávam sa, že takáto predstava replikátora sa od mystických predstáv v Biblii principiálne nelíši, pretože sa nijako nedozvedáme ako sa z neživej hmoty riadenej čisto fyzikálnymi zákonmi vynorí hmota živá, nadaná prinajmenšom novými (fyzikálnymi zákonitosťami nevyjadriteľnými) vlastnosťami: sebareplikovateľnosťou a schopnosťou ovplyvňovať pravdepodobnostný charakter svojho osudu. Skrátka Boh je ten ktorý je.
Nech je ako chce, označme túto predstavu replikátora aktívny replikátor . Pre ďalší text považujem za podstatné uviesť podstatnú vlastnosť aktívneho replikátora: aby mohol byť aktívny, musí mať možnoť rozhodovať sa, musí mať zodpovedajúce informácie o prostredí, ale hlavne všetky zodpovedajúce informácie o replikácii.
Na inom mieste Dawkins uvádza:
… Další, složitější možnost je, že stavební jednotky nemají afinitu vůči svému druhu, ale vůči určitému jinému druhu stavební jednotky. Pak by replikátor nebyl předlohou pro stejný řetězec, ale pro řetězec „negativní“, komplementární; podle toho by se pak zase vytvořil původní, pozitivní. (Richard Dawkins: Sobecký gen, kapitola Replikátor)
Ak máme brať replikátor ako šablónu , tak je ovšem situácia iná. Replikátor je iba informácia, ktorú niečo interpretuje . Stáva sa pasívnou súčasťou procesu. V tom prípade ovšem časť informácie o replikácii je mimo samotný replikátor a časť je v ňom samom. A skutočne to aj sám Dawkins takto popisuje. Ak má nejaká molekula afinitu k replikátoru tak je to ich spoločná vlastnosť, nie iba vlastnosť replikátora. Ešte zreteľnejšie je to v príklade "negatívnej" komplemenarity. Tam je replikátor informácia, ktorá má afinitu voči niečomu opačnému a z toho by vznikol až rovnakým procesom orginál.
Vidíme teda, že v tejto predstave replikátor vystupuje ako replikovaná informácia, teda pasívne. Nazveme takýto replikátor pasívnym replikátorom .
Tak ako pán Dawkins… aktívny, či pasívny replikátor?
Kto čítal knihy Richarda Dawkinsa, tak vie, že celá jeho predstava o evolúcii je založená na predstave aktívneho replikátora. Gén je určujúci, má vlastné záujmy, všetky biologické štruktúry ako proteíny, bunky, organizmy, sú iba pasívne vehikly aktívneho génu. Dokonca aj mém je aktívny replikátor, neraz má organomorfné vlastnosti, je vnímaný ako vírus, nákaza a podobne.
Situácia tak „aktívne“ nevyzerá, ak chce Dawkins popísať podstatu replikátora v reálnom živote. V Slepom hodinárovi mu prisudzuje tri „ingrediencie“, ktoré by mali popísať jeho podstatu:
- sebareplikovateľnosť,
- chybovosť replikácie,
- a už spomínanú moc ovplyvňovať svoj osud.
Pozrime sa na ním uvedený reálny dej replikácie:
Vidíme, že aktívna RNA Qbeta vírusu potrebuje na svoju replikáciu nevyhnutne mechanizmy bunky Escherichia coli, inak by sa vôbec nenamáhal vniknúť do nej. Bunka mu totiž musí vyrobiť množstvo amínokyselín a iných stavebných prvkov, aby mala z čoho vyrobiť RNAreplikázu, samotné RNA molekuly a zopár bielkovín, ktoré potrebuje na výrobu obalov, v ktorých sa novovzniknuté RNA prepravujú k ďalším infikáciám.
Celý dej však môžeme popísať aj opačne. E-coli aktívne rozmnožuje RNA z Qbeta vírusu tak, ako rozmnožuje aj iné vlastné štruktúry. Navyše to, či sa Qbeta rozmnoží, alebo nerozmnoží, závisí od toho, či iné aktívne deje (pohyb tekutého prostredia, v ktorom vírus je, alebo pohyb obsahu čriev, kde sú tak E-coli, ako aj Qbeta „unášané“) dopravia replikátor z Qbeta k príjemcovi E-coli. A ani to nestačí, E-coli musí dostať Qbeta cez bunkovú membránu dnu, musí vytvoriť RNAreplikázu a mnoho iných dejov, ktoré sú nevyhnutné, aby sa RNA z Qbeta mohla údajne sebareplikovať. Je to iba iný uhol pohľadu a neviem prečo by nemal byť rovnako oprávnený. A to ani nehovoriac o tom, že celá moc ovplyvňovať svoj osud sa rozplýva v takom obrovskom množstve zásahov z prostredia, že je iluzórne predpokladať akúkoľvek „intencionalitu“ RNA Qbeta voči svojmu prostrediu bez toho, aby sme predpokladali rovnakú intencionalitu prostredia voči tejto RNA.
Ale povedzme, že to nie je až taký dobrý príklad, pretože ako RNA, tak aj Echerichia coli sú už príliš zložitý systém na to, aby sme mohli demonštrovať najmä sebareplikovateľnosť a moc ovplyvňovať svoj osud. Preto skúsme pokračovať iným príkladom:
V každom prípade je zrazu miesto RNA ako aktívneho replikátora aktívnou sústava RNA-RNAreplikáza. Z RNA je zrazu šablóna , ktorá je replikovaná RNAreplikázou, A šup — odrazu sú tu aj potrebné látky , ktoré boli do procesu pridané Solom Spiegelmanom a jeho kolegami.
Po tom, ako popisuje túto evolúciu v skúmavke, kde sa týmto vedcom podarilo demonštrovať prírodný výber skoro v priamom prenose a ďalšie podobné experimenty, Dawkins konštatuje:
Zasa ide o replikačné stroje, ktoré vyrábajú replikátory (RNA), RNA je teda pasívna šablóna a nie naopak... a vlastne akože nie? Veď tie replikačné stroje sú vyrábané podľa nejakej inej RNA, či DNA! To však nebráni Dawkinsovi vyjadriť len o odstavec ďalej:
Na dnešních organismech je máme stále na očích. Jsou to oči, kůže, kosti, prsty, mozky a instinkty. To vše jsou nástroje k replikaci DNA. Jsou výsledkem působení DNA v tom smyslu, že rozdíly v očích, kůži, kostech, instinktech atd. jsou vyvolány rozdíly v DNA. Mají vliv na replikaci DNA která je způsobila tím, že ovlivňují přežití a rozmnožení svých těl, jež obsahují tutéž DNA, a proto sdíle jí její osud. Proto DNA sama ovlivňuje svou vlastní replikaci prostřednictvím vlastností těl. Můžeme tedy uzavřít, že DNA má moc nad svým vlastním osudem. Těla, jejich orgány a způsoby chování jsou nástroje používané touto mocí.
Když mluvíme o moci, máme na mysli to, jak replikátory ať už sebevíce nepřímo ovlivňují svou vlastní budoucnost. Nezáleží na tom, kolik článků má řetězec příčin a následků. Jeli příčinou sebe replikující věc, mohou být následky předmětem přírodního výbě ru.Opäť sa dostávame k hypotetickým aktívnym vlastnostiam replikátora, ktorý je zasa z ničoho-nič umiestnený v molekule DNA, či RNA. A nielen to. Tieto hypotetické vlastnosti, vyvolávajú kauzálny reťazec z ďalších hypotetických vlastností a tak ďalej, až po celý organizmus. Stačí zmena písmena v géne na DNA… a všetko sa údajne zmení, pretože tam je zdroj všetkého.
Musím pripomenúť — je priveľa argumentov na to, že takto to v organizmoch skrátka nefunguje, ako to je v skutočnosti si možno prečítať napríklad v knihe Antona Markoša Povstávání živého tvaru .
Pre projekt Anima Et Machina je podstatné to, že neurčitá a voluntaristická predstava replikátora viedla k predstave mému — aktívneho replikátora, ako analógiu génu, takisto chápaného ako aktívnu entitu. Domnievam sa, že toto malo pre memetiku závažné dôsledky, o ktorých (dúfam) ešte bude reč.
Tak ako pán Dawkins: aktívne, či pasívne? Vždy keď Dawkins popisuje replikátor ako princíp, tak hovorí o aktívnom replikátore. Tento má vlastnú intencionalitu a všetko ostatné sú vehikle, prostredníctvom ktorých túto intencionalitu realizuje. Lenže Dawkins je zároveň biológ a pri popise fungovania replikátora ako biologickej štruktúry je jeho replikátor pasívny. Obávam sa, že táto otázka je kľúčová, a to ani nehovoriac o tom, že samotný koncept replikátora je takto spochybniteľný.
Áno, či nie?
Ako sme videli v predchádzajúcich
kapitolách, Richard Dawkins sa pokúsil vyjadriť podstatu evolúcie života
ako evolúcie informácie uloženej v molekule DNA, či RNA. Podstatou bola
informácia, nie samotný molekulárny nosič tejto informácie. čo je na
profesora z Oxfordu pomerne prirodzený záver. Túto informáciu nazval
replikátorom.
Zhrňme si základné vlastnosti replikátora, tak ako ho Dawkins prezentuje.
- Celistvosť a ohraničenosť — replikátor je do seba uzatvorený kus informácie, ktorý svojou vnútornou skladbou nesúvisí s prostredím, v ktorom sa vyskytuje. Dokonca môže prechádzať cez rôzne nosiče, ale sám osebe po týchto prechodoch ostáva nezmenený. Viď kopírovanie génu z DNA na RNA.
-
Stabilita a nestabilta štruktúry — je vlastne
základnou hybnou silou evolúcie. Kým na jednej strane si replikátor
udržuje stabilitu svojho usporiadania neustálym replikovaním, toto
replikovanie nie vždy prebehne presne, čím vznikajú odchýlky, ktoré sú
podrobované tlaku prírodného výberu. Úspešnejšia odchýlka prežije na
úkor menej úspešnej.
- Intencionalita, účelnosť — replikátor sa prosto iba nevyskytuje, ale vyskytuje sa a koná vždy za nejakým účelom. Napokon to má byť jeho základná rozlišovacia charakteristika od neživej prírody. Kým kryštál má svoju stabilitu v čase danú akosi „samú od seba“, replikátor je vysoko nestabilný a stabilitu dosahuje tak že sa neustále replikuje, teda inak povedané: replikátor sa neustále replikuje za tým účelom, aby sa zachovala jeho štruktúra, teda informácia. Replikátor sa spája do DNA s inými replikátormi za tým účelom, že spolu majú väčšiu šancu na replikáciu svojich štruktúr. A tak ďalej… replikátoy sa obalili bunkovymi membránami za tým účelom, aby ľahšie udržali svoju štruktúru… replikátory si vytvorili svaly, nervový systém, telá, a pod… za tým účelom, aby sa mohli rozmnožiť do čo najviac kópií, a tým si udržali stabilitu svojej štruktúry.
Z uvedeného pre nás vyplývajú, podľa mojej mienky dve otázky, ktoré sa pokúsim následne rozobrať.
Tušenia symbióz
Stabilita pojmov môže byť frustrujúca. Ak dnes začíname opatrne
hľadať hlbší význam obrovského množstva poznatkov, ktoré sa nakopili
hlavne s vývojom vied, tak sa neraz ocitneme v situácii, keď na seba
narážajú do očí bijúce pojmy s rovnakou štruktúrou, ale s iným
historickým pozadím, prípadne iným koncepčným zázemím. Roztrieštenosť
poznania v rôznych vedeckých disciplínach, teológiách či filozofiách
prakticky znemožňuje v stromoch vidieť les. Obsiahnutie toho lesa,
dôsledku predpokladu, že bytie je jednotné a nedeliteľné, vyžaduje
nielen poznanie monohých zdanlivo vzájomne nesúvisiacich poznatkov, ale
aj istú vzburu proti pojmom. Pojmy (majúce svoju štruktúru danú práve
tou roztrieštenosťou) sa stali z nevyhnutného komunikačného nástroja
medzi sapientami brzdou uchopovania štruktúry prostredia. Jedným z
takýchto pojmov je podľa mojej mienky aj symbióza, rep.symbiotizmus.
Slovo symbiotizmus použil v roku 1877 Albert Bernhard Frank aby dáko vyjadril fakt, že sú také orgsnizmy, ktoré pre svoj život potrebujú iné organizmy. Za každým slovom je pojem, teda nejaká štruktúra modelu prostredia, ktorá sa vytvára v našich hlavách. Za symbiotizmom (javom spolu-žitia) Frank zjavne videl zreteľné ukážky spolu-žitia dvoch druhov organizmov tak tesného, že ho nemožno oddeliť. Súžitie lišajníkov s hubami, parazitov a hosteľa, mravcov a vošiek, rastlín a ich opeľovačov… Nemám dosť elánu na to, aby som skúmal, čo viedlo Franka k popisu spolužitia organizmov, nijako by ma však neprekvapilo, ak by to nebol aj dôsledok Darwinovho pohľadu na evolúciu, v ktorom koncepcia prírodného výberu zdôrazňuje vzťah medzi organizmom a jeho prostredím.
Pojem symbióza je od tých čias na svete a žije si svojím životom. V prvom rade sám seba ohraničil
definíciou
,
čím pri svojej replikácii do ďalších hláv vytvára pocit vylúčenosti a
výlučnosti. Ak dnes povieme: symbióza, každý si predstaví nejakú tú
formu nerozlučného spolužitia dvoch organizmov. Ak sa pozrie na
definíciu trebárs vo
Wikipédii
, nanajvýš ak vyjadrí počudovanie, že za symbiózu biológovia považujú aj parazitizmus.
Biológovia už z povahy vlastnej tradície jav symbiózy pekne
zaklasifikovali, roztriedili na druhy a spôsoby. Všetko vyzerá byť
úhľadné a vcelku rozumné, ako keby nebolo potrebné k veci uviesť nič
podstatné.
Lenže ja s tým mám problém, ktorý sa dá zhrnúť do prostej otázky: prečo je symbióza, resp. symbiotizmus? Tí istí biológovia, ktorí považujú za hybný moment života gén uložený v molekule DNA, gén, ktorý má byť alfou a omegou všetkého živého, gén, ktorý je tou elementárnou jednotkou prírodného výberu, a preto je vlastne jediným skutočným subjektom života a ním vytvorené bunky a telá sú iba vyjadrením tejto subjektivity, teda tí istí biológovia, ktorí vychádzajú zo subjektivity génu zrazu nemajú problém so systémom dvoch organizmov, ktorých spolužitie je sotva subjektívnym vyjadrením jedného spoločného génu (prípadne niekoľkých spoločných génov).
S odretými ušami si viem predstaviť, že nejaký spoločný gén určuje
spolužitie mravca a vošky. Spolužitie vyzerá tak, že mravec nosí voške
potravu a chráni ju pred predátorom a ona mu za to vylučuje potravu, z
ktorej mravec žije. Dobre. Nech teda je nejaký gén, ktorého reprodukčný
úspech (
fitness
)
sa zvýši ak je vo voške a súčasne v mravcovi tak, že usmerní ich
správanie k symbiotickej konformácii. Ale predpokladať nejakú podobnú
génovú konštrukciu u symbiózy včely a kvetu by bolo zrejme riadne
pritiahnuté za vlasy. Taký gén by musel byť riadne starý, pretože by
musel byť už v organizmoch, ktoré predchádzali rastlinám a článkonožcom,
musel by byť teda už v jednobunkových predchodcoch. Na čo by tam
vlastne slúžil?
Vyzerá to teda skôr tak, že dôvod pre symbiotizmus treba hľadať mimo génocentrickú paradigmu súčasnej biológie. Z pohľadu biológie je symbiotizmus diktovaný prostredím, je to niečo ako vzťah predátora a koristi, či iné ekologické zákonitosti, ktoré vytvárajú selekčný tlak na taký výber génov, ktoré symbiotizmus umožnia. A tu pociťujem rovnakú podivnú koncepčnú dichotómiu medzi všetko určujúcim génom a neurčitým definovaním prostredia a jeho zákonitostí ako vo viaxerých biologických pojmoch.
Napríklad taká otázka vzťahu génotypu a fenotypu, teda vzťah medzi súborom génov organizmu a súborom znakov, ktoré vlastne tvoria to fýzis, telo organizmu. Dnes prevláda viera, že gény určujú znaky organizmu. V súvislosti so symbiózou je potom na mieste položiť si otázky:
- Je v symbiotickom vzťahu symbiont znakom organizmu? Teda napríklad je pre slnečnicu včela jej fenotypovým znakom? Je pre črevnú baktériu sapient jej fenotypovým znakom? Ak gén určuje správanie sa organizmu, malo by tomu tak byť.
-
Ak považujeme vtáčie hniezdo, mravenisko mravcov, úľ včiel, vodnú
hrádzu bobrov, či jazyk sapienta za tzv rozšírený fenotypový znak, prečo
to isté neplatí v prípade symbiózy? Je teda včela rozšíreným
fenotypovým znakom fialky?
- Ak je symbióza fenotypovým znakom, ako je možné, že sa vyskytuje u najrôznejších druhov organizmov? Je tu síce známy jav, kedy rovnaké podmienky v prostredí vytvoria tlak na rovnaké znaky organizmu. Napríklad plutvy majú aj ryby ale aj cicavce — delfíny. Lenže v tejto súvislosti ostáva do génostrednej predstavy nezapadajúci fakt, že plutvovitosť je vlastnosťou prostredia, ktorá sa replikuje na všetky vhodné organizmy. Vlastnosť, ktorá si vyberá vhodné gény, nie naopak. Podobne to je aj so symbiotizmom, ako vlastnosťou prostredia, ktorá sa replikuje na vhodné organizmy a určuje selekciu ich génov. Inými slovami: znaky organizmu určujú vhodné gény, nie naopak. Znaky sú tie subjekty evolúcie a gény sú ich objektami.
Naozaj, prečo neskúsiť pohľad na symbiózu, ako na znak, ktorý má
vlastnú subjektivitu, teda sa rozmnožuje na organizmoch vždy, keď je to
možné. Prečo by mal byť jediným replikujúcim sa subjektom iba gén a
nemôže ním byť aj vlastnosť prostredia, či kooperácia organizmov? Aj
jeden z propagátorov „sebeckosti“ génov, Richard Dawkins nás nabáda, aby
sme gén chápali ako informáciu, nie ako molekulu. No a čo iné je
informácia, ako nejalé usporiadanie prvkov do celku — znaku. Potom je
symbiotické spolužitie organizmov tak isto znakom, ako trebárs šupiny,
či klepetá. Znak, teda usporiadanie, teda informácia sú to, čo je
replikované.
Samozrejme znakom môže byť aj kúsok DNA, ktorý je neustále
replikovaný bunkou. Podľa niektorých biologických koncepcií si kúsky
molekuly RNA vytvorili DNA na to, aby si mali kam uložiť informáciu pre
vlastnú replikáciu. Na organický svet môžeme nazerať ako na súbor
znakov, ktoré sú v rôznej miere, s rôznou úspešnosťou replikované. Prečo
by sme mali myšlienkovo vydeľovať organizmy z ich prostredia? Na jednej
strane uznávame, že organizmy tvoria ekosystém, ktorý následne vytvára
selekčný tlak na výber vlastností (znakov) organizmov, súčasne však máme
tendenciu oddeľovať vlastnosti ekosystému (napríklad symbiotizmus,
potravinová hierarchia, populačné zákonitosti atď) od vlastností
organizmov.
Vidím v tom systémovú nekonzistentnosť. Ako príklad uvediem znova symbiotizmus. Ak je chápaný ako tesné súžitie dvoch organizmov (viď definícia vo Wikipédii), tak sa táto definícia vzápätí oslabí príkladom symbiotizmu: eukaryotická bunka. Eukaryotické bunky, z ktorých sme poskladaní), obsahuje okrem DNA a nejakých molekulových strojčekov aj mitochondrie, ktoré majú vlastnú DNA. Mitochondrie slúžia našim bunkám ako zdroje energie. Zdá sa, že to boli pôvodne samostatné bunky, ktoré sa v evolučnej minulosti stali súčasťou dnešných eukaryotických buniek. Žijú s nimi v symbióze. Podstatné pre naše úvahy však je, že samé sa nedokážu rozmnožiť, stratili časť svojej DNA, vlastné rozmnoženie nechávajú na materskú bunku. To už nevyzerá na „čisté“ súžitie dvoch samostatných organizmov, ktoré by dokázali prežiť osve. No ale to nedokáže ani väčšina génov uložených v DNA. Prečo potom molekulu DNA nepovažovať za symbiotické súžitie génov daného organizmu?
Čo je vlastne podstatou predstavy symbiotizmu? Pôvodne z pohľadu Alberta Bernharda Franka zrejme výhodnosť pre symbionty. Dnes však z pre mňa nepochopiteľných dôvodov definícia symbiózy hovorí o tesnom súžití organizmov. V čom je to tesné súžitie, v skutočnosti, že minimálne jeden zo symbiontov neprežije bez toho druhého? V takom prípade je táto definícia dosť vágna a nepresvedčivá, pretože prísne vzaté — všetci tvoríme spolu ekosystém a v rôznej miere sme na sebe závislí. Potom je „tesnosť“ subjektívny pocit hodnotiaceho biológa, alebo normatívne určená? Ak áno, tak postráda zmysel. Načo je taká biologická definícia, ktorá má povahu normatívneho aktu? Napríklad ak sa živíme poľnohospodárskou produkciou, ktorej základom sú rastliny, tak sotva prežijeme bez pôdnych baktérií. Žijeme teda v ako tesnom symbiotickom vzťahu s pôdnymi baktériami?
Skúsme sa na vec pozrieť z iného zorného uhlu, ako je génocentrický,
len si z neho skúsme zachovať základnú myšlienku, že podstatou génu nie
je molekula DNA, ale to, že je to replikovaná informácia, pričom
informácia je usporiadanie niečoho. Ak je gén informácia, tak je
usporiadaním molekúl v DNA makromolekule. Toto usporiadanie je
replikované ako celok, teda celý jeden kúsok DNA sa rozmnožuje v tom
usporiadaní molekúl, ktoré tvoria gén. Replikáciu zabezpečuje prostredie
bunky. Pre gény je bunka ich prostredím.
Pozrime sa na symbiózu z rovnakého pohľadu. Spolužitie symbiontov môžeme vyjadriť ako ich určité usporiadanie, ako informáciu, ktorá je prostredím, teda biotopom replikovaná ako jeden celok. V takomto zmysle sú akékoľvek vzťahy organizmov v biotope planéty rovnakej kvality. Vzťah oeganizmov, teda ich usporiadanie, sú informáciami, ktoré sú replikované v ich štruktúre ako celok. Vzťah predátora a koristi, hoci je replikovaný na rôzne konkrétne typy organizmov, je vo svojej štruktúre rovnaký a vždy, keď sa v biotope vytvoria vhodné podmienky, tak sa vzťah predátora a koristi zreplikuje na ďalšie organizmy. Podobne aj vzťah parazita a hostiteľa, vzájomne výhodný vzťah symbiontov, či vzťah potravinového reťazca. Skrátka akýkoľvek vzťah medzi organizmami.
Takýto pohľad je dôsledným akceptovaním súčasnej predstavy
biologickej evolúcie, ktorá chápe gén ako replikovanú informáciu, nie
ako molekulu. Ten istý pohľad však zároveň nepriznáva génu výsadné
postavenie ako jediného objektu prírodného výberu. Nevidím pre to
najmenší dôvod. Ak by mal byť gén jediným objektom prírodného výberu,
tak by bolo treba túto výlučnosť zdôvodniť a tiež by bolo nevyhnutné
vysvetliť nielen tvarovú stálosť génov, ale aj tvarovú stálosť
ekologických vzťahov (napríklad aj symbiózy) a ich opakovanosť,
replikovanosť.
Evolučná biológia napríklad popisuje jav mutačných ťahov a tlakov. Tento jav vyjadruje skutočnosť, že určitý typ mutácie je pravdepodobnejší ako iný, čisto náhodný. Kým čisto náhodné mutácie génov môžu byť prírodným výberom eliminované, iné ktoré vznikajú buď z fyzikálneho usporiadania molekuly DNA, alebo z daného prostredia v bunke vznikajú v určitom percente neustále, aj keď sú prirodzeným výberom eliminované. Prečo podobne nenazerať na symbiotický vzťah ako tlak ekosystému na vznik určitého typu spolužitia vždy, keď je to možné?
Inými slovami: prečo nepredpokladať, že evolučné zákonitosti neplatia iba pre organizmy, tak ako ich prvý raz definoval Ch. Darwin, ale aj pre vzťahy medzi organizmami?
Akýkoľvek vzťah medzi organizmami je ich usporiadanie do nejakej štruktúry. Je to informácia. Akékoľvek usporiadanie molekúl v DNA môžeme rovnako chápať ako ich vzťah, ako informáciu. A tiež akoko ich ekosystém. Niektoré usporiadanie je stabilnejšie, iné menej, niektoré sa ľahšie replikuje iné menej. V oboch prípadoch sa teda uplatňuje tvorivá sila prírodného výberu. A v oboch prípadoch sa uplatňuje vytváranie stability usporiadania, ako zákonitý výsledok prírodného výberu.
Napokon, prírodný výber vždy bol a je o vzájomnom vzťahu organizmov, o
ich usporiadaní a ich usporiadavaní. Prírodný výber je tvorivý akt, keď
sa nejaké usporiadanie organizmov vo vzájomnom vzťahu znovu usporiada v
novej reinterpretácii týchto vzťahov. Pokiaľ to uznávame vrámci jedného
druhu organizmov, či dokonca v kontexte súťaže jednotlivých génov,
nevidím najmenší dôvod pre to, aby sme to isté nemohli chápať vo
vzťahoch medzi druhmi organizmov.
A rovnako nevidím principiálnu prekážku pre to, aby sme také druhy vzťahov, ako je symbióza, parazitizmus, potravinová hierarchia, frekvenčne závislá selekcia, či evolučne stabilné stratégie a iné zákonitosti, ktoré sme identifikovali v evolučnej biológii a ekológii, nemohli skúšať hľadať aj v iných druhoch usporiadaní, dokonca aj tam, kde by sme ich doteraz nehľadali. Opäť sa inšpirujme symbiotickým vzťahom, len si ho prestaňme spájať s organizmami. V ďalšom texte budem pod symbiózou chápať iba ten druh symbiotického vzťahu, kde z neho majú obe strany prospech, nech už je tým prospechom čokoľvek, pričom tento vzťah je replikovaný, rozmnožovaný (keď už sa vytvoril) ako jeden celok.
Po viacerých pokusoch vysvetliť fenomén kutúry sa až v poslednej tretine 20. storočia objavila skutočne darwinistická koncepcia, ktorú jej autor Richard Dawkins a jeho nasledovníci nazvali teóriou mémov. Teória sama osebe trpí mnohými ťažkosťami a koncepčnými nehomogenitami. Ak však chceme zostať darwinistami, nemáme k teórii mémov alternatívu. V teórii mémov chápeme mém ako kultúrny pandat génu. V oboch prípadoch ide o podstatu, teda o informáciu, usporiadanie.
V prípade kultúry usporiadanie čoho? Na tejto otázke dnes zatiaľ
teória mémov stroskotáva. Príčinou je podľa mojej mienky
kontraproduktívne hľadanie materiálneho nosiča mému, pričom sa neustále
poukazuje na to, že gén má svoj materiálny nosič — molekuly DNA a RNA.
Zabúda sa pri tom na to, že informácia je nemateriálna v zmysle
samotného nosiča informácie, nosiča, ktorý nie je bytostnou podstatou
informácie. Informácia je in-formácia, teda usporiadanie niečoho do
formy. Ak má byť teória mémov k nejakému prospechu, tak musí v prvom
rade identifikovať, čoho formovaním je mém. Tak ako som v prípade
symbiózy organizmov ponúkol koncepciu znaku, ako replikovanej
informácie, tak pokojne môžem v prípade kultúry urobiť to isté. Mém je
znak kultúry, ktorý je replikovaný. Kým replikovaný? Jednoznačne
spoločnosťou sapientov. Spoločnosť je možné chápať ako ekosystém
spoločenských interakcií. Tieto interakcie sú evolučným produktom,
ktorého zmyslom bolo (tak ako aj v iných súvislostiach) zabezpečiť
prežitie génov, znakov, jedincov, či druhov, alebo dokonca
ekolosystémových interakcií.
Každá takáto interakcia je znakom, ktorý môžeme nazvať mémom. Interakcia medzi organizmami je v súčasnej biológii chápaná ako fenotypový znak, ktorý je vehiklom nejakého génu, alebo génov. Také jednoduché to však nemôže byť. Ak sa veci prizrieme bližšie, tak sa ukáže iný obraz. Existuje viacero foriem pohlavného výberu, ktorý môžeme pokojne chápať ako jednu z viacerých oblastí sociálnej interakcie. Kým jedny druhy uplatňujú čisto pohlavný výber samičí, iné zasa vzájomný pohlavný výber samcov a samíc. V prvom prípade je starostlivosť o potomstvo buď nulová, alebo nízka, zabezpečená samicami. V druhom prípade sú obyčajne potrebné vysoké rodičovské investície samca a samice. To, ktorá forma sociálnej interakcie sa presadí závisí od životných podmienok prinajmenšom rovnako, ako od génov, ktoré formujú fenotypový znak sociálnej interakcie. Dá sa povedať, že prostredie replikuje tie gény, ktoré sú pre prežitie v ňom najvýhodnejšie. Replikuje sa pri tom znak — mém — sociálnej interakcie. Mém v takomto ponímaní nie je (na rozdiel od definície R. Dawkinsa a jeho nasledovateľov) iba dôsledok napodobňovania výhodného správania iných č lenov tlupy, ale oveľa fundamentálnejší sociány jav, je to akýkoľvek znak sociálnej interakcie.
Toto vnímanie mému umožňuje chápať kultúru ultimatívne darwinisticky ako ekosystém mémov. A ako v každom ekosystéme, aj tu môžeme identifikovať výhodné spolužitia — symbiózy. Dokonca môžeme povedať, že symbiotická interakcia je základnou interakciou v mémickom ekosystéme. Veď čo iné je mémplex (komplex mémov) ako výhodné spolužitie — symbióza — mémov? A nielen to. Mémplexy tak isto dokážu vytvárať symbiózy. Napríklad mémplex peňazí dnes vytvára veľmi výhodné symbiózy s rôznymi druhmi mémplexov moci. Mémplex moci tým získava od sapientov potvrdenie výlučnej replikácie určitej konkrétnej formy spoločenského usporiadania, mémplexy peňazí získavajú sapientom vnútenú formu ich replikácie v nekomoditnej, teda dlhovej forme, čo im umožňuje vytvorenie oveľa viac kópií ako v komoditnej forme.
Symbióza ako výhodnosť určitého usporiadania entít môže byť prejavom nejakej základnej vlastnosti Vesmíru. Ak chceme, prečo by sme sa napríklad nepozerali na každý atóm ako na výhodné usporiadanie častíc s ich konkrétnymi kvantovými stavmi a s daným stavom prostredia, v tomto prípade určenom hustotou energie? Alebo usporiadanie kvarkov do bariónov ako sú protóny, alebo mezóny? Kvantové čísla môžeme chápať v takomto kontexte ako znaky, ktoré spoluvytvárajú usporiadanie častíc.
Napokon ak pripúšťame evolúciu v organickom svete, mali by sme z
prepokladu jednoty a nedeliteľnosti bytia nejako k organickej evolúcii
vo vyvíjacom sa Vesmíre dospieť ako k vývojovému dôsledku
predchádzajúceho vývoja. Evolúcia by mala byť základnou vlastnosťou
Vesmíru. Dnes stále nemáme fyzikálnu teóriu, ktorá by vedela vysvetliť
ako prejsť od termodynamických zákonov k zákonom usporiadavania
termodynamických systémov do štruktúr. Formy usporiadavania známe ako
darwinizmus viac-menej uznávame pre organické termodynamické štruktúry a
možno pre spoločenské interakcie. Tušíme, že musia byť dôsledkom
nejakých obecnejších evolučných zákonitostí.
Symbiotizmus je možno jednou z nich.